ponedjeljak, 30. rujna 2019.
nedjelja, 8. rujna 2019.
Uređenje postova
Još treba da uredim postove što sam sad postavio.
Uređeni su postovi od zadnjeg do članak o baraminologiji, zatim članak o datiranju K-Ar je uređen. Ostali noviji nisu još uređeni.
Uređeni su postovi od zadnjeg do članak o baraminologiji, zatim članak o datiranju K-Ar je uređen. Ostali noviji nisu još uređeni.
Neuspeh „konkordizma“ dugog doba
str. 57-59, the Genesis account, teološki, istorijski i naučni komentar na Postanje 1-11,
Džonatan Sarfati, PHD, F.M.
Zapazi da okvir gledište ne pokušava da
uklopi dane Postanja sa pravim istorijskim danima. Ali, ima onih koji
žele da rastegnu dane i naivno misle da se oni mogu uklopiti sa redom
Stvaranja, ako već ne sa vremenskim okvirom. Ovo je tip „konkordskog“
gledišta.
Međutim, postoje glavne kontradikcije
između doslovnog čitanja Pisma i reda tvrđenog od
uniformacijske/evolucione „nauke“. Ovo je objašnjeno u Tablici 2.2. Neki
od glavnih naglasaka su:
2.
Evolucija postulira da su prvi živi organizmi bili jednoćeliski
organizmi koji su proizašli iz okeanske iskonske supe hemijskom
evolucijom.3 Nakon toga živa bića su evoluirala u morima mnogo prije kopnenih biljaka i životinja i još ranije od drveća. Ali, Postanje otkriva da je Bog stvorio kopnene biljke, uključujući drveće prvo.
3. Evolucija uči da su ihtiosaurusi i drugi morski gmizavci
evoluirali od kopnenih gmizavaca i da su kitovi evoluirali od kopnenih
sisavaca, koji su evoluirali od drugih kopnenih gmizavaca. Dan-dobci
odano tvrde da su prethodni stvoreni prije narednih. Slično tome,
evolucionisti veruju da su ptice i pterosaurusi evoluirali od kopnenih
reptila dok su šišmiši evoluirali od kopnenih sisavaca. Međutim,
Postanje izričito uči da je Bog stvorio morska i leteća stvorenja 5.
dana, dan prije nego što je stvorio kopnena stvorenja, 6. dana. Ovo je dovelo do vrlo maštovitog izvrtanja Pisma da se eon-„dani“ preklapaju, a ne da slede jedan iza drugog.4
4. Svaki oblik evolucije mora čvrsto poreći doslovno prvog
pojedinačnog muškarca i ženu koji su jedini predci svih drugih ljudi
koji su ikada postojali.5 To jest, oni kažu da je populacija majmunolikih stvorenja evlouirala u populaciju
ljudi. Postanje s druge strane uči da je prvi čovek stvoren ne od živih
stvorenja, nego od nežive materije (praha zemljskog), koja je postala
živa kada je Bog udahnuo u nju (Postanje 2:7). I, prva žena slično tome
nije imala majku, nego je stvorena od čovečijeg rebra (Postanje
2:21-24).
Kreacionisti dan-dobci su prošli malo
bolje, oni slepo prihvaćaju evoluciono datiranje. Evolucionisti datiraju
fosil nesumnjiivo Homo sapiensa na otprilike 200.000 godina,
mnogo ranije nego što Adam može biti čak sa najapsurdnijim rastezanjem
biblijskog vremenskog okvira danog u Postanju 5 i 11. Ovo je dalje
objašnjeno u „Ljudska smrt-veliki problem za milione godina.“, Poglavlje
13.
5. Sva gledišta-milijardi godina stavljaju smrt prije greha. Teško
je dovoljno naglasiti važnost ovoga. Ova gledišta uče da čim su se živa
bića pojavila ona su takođe i umirala. Međutim, prva zabeležena smrt biblijskog živog stvorenja (hebrejski nefeš haja 5) se desila nakon što su Adam i Eva zgrešili,
kada je Bog ubio životinju da napravi kožnu odeću za njih. Zato je
široka sekcija ovog komentara (knjige) posvećena poreklu smrti od greha
(vidi „Fizička smrt“, pogl. 13).
Tablica 2.2.: Kontradikcije između Postanja i uniformacijske „nauke“ u sledu događaja.
| Dan | Biblijsko stvaranje | Veliki prasak, Evolucija | Evolucioni sled | |
| 1a | Nebo i Zemlja | Svetlost | 1 | |
| 1a | Tama | Prostranstvo (expanse) | 2 | |
| 1a | Voda, okeani | Zvezde | 3 | |
| 1b | Svetlost | Voda | 4 | |
| 2 | Prostranstvo (expanse) | Sunce | 5 | |
| 3a | Suva zemlja | Zemlja i solarni sistem | 6 | |
| 3b | Prvi život: kopnene biljke, drveće | Suva zemlja | 7 | |
| 4 | Sunce, Mesec, zvezde | Okeani | 8 | |
| 5 | Ribe | Prvi život: jednoćelijski organizmi | 9 | |
| 5 | Kitovi (uključujući delfine) | Smrt | 10 | |
| 5 | Ihtiosaurusi, mosaurusi | Ribe | 11 | |
| 5 | Pleziosaurusi, pliosaurusi | Drveće | 12 | |
| 5 | Ptice, šišmiši, pterosaurusi | Kopneni gmizavci | 13 | |
| 6a | Kopni gmizavci („životinje što se miču“) | Ihtiosaurusi, mosasaurusi, pleziosaurusi, pliosaurusi, pterosaurusi od kopnenih gmizavaca | 14 | |
| 6a | Kopneni sisavci | Kopneni sisavci (od kopnenih gmizavaca) | 15 | |
| 6b | Jedan čovek od praha | Ptice od kopnenih gmizavaca | 16 | |
| 6c | Jedna žena od čovečijeg rebra | Šišmiši i kitovi (od kopnenih gmizavaca) | 17 | |
| Nekoliko dana nakon nedelje Stvaranja | Smrt (zbog Adamovog greha) | Populacija majmunolikih stvorenja je evoluirala u populaciju ljudi | 18 |
1. Henry.J., The elements of the universe point to creation: Introduction
to a critique of nucleosynthesis theory, J.Creation 20(2):53-60,2006.
2. Sarfati,J., Solar system origin: Nebular hypothesis, Creation 32(3):34-35, 2010, creation.com/nebular.
3. Batten, D., Origin of life: An explanation of what is needed for
abiogenesis (or biopoiesis), creation.com/origin-of life, 26. novembar
2013.
Još jedna nogata zmija?
Šta kreacionisti da misle o tome?
Džonatan Sarfati
Objavljeno: 29. aprila 2008. godine
Evolucinisti su uzbuđeni zbog nedavnog otkrića, stražnjih nogu na fosilu zmije.1
Ali, šta je zapravo pronađeno i koja su prihvatljiva i neprihvatljiva objašnjenja?
Šta je pronađeno?

Slika: news.bbc.co.uk
Zapravo, sam fosil je prilično stara
vest. Francuski paleontolog Žan Klod Raž i Fransoa Escvil su ga otkrili
2000. godine otisnutog na tankom krečnjačkom bloku blizu Lebanskog sela
al-Namura. 85 cm (33,5 in) dugačko stvorenje je uvršteno uniformacijskom
stratigrafijom Cenomijskoj fazi, prvoj fazi gornjeg Kreatocena i
„datirano“ je na oko 94 miliona godina starosti. Prvi put je stvorenje
nazvano Podophis descousensi,2 ali ime roda je otklonjeno, tako da je
preimenovano u Eupodophis.3
Aleksandra Houssaye iz Nacionalnog muzeja
prirodne istorije u Parizu i njen tim su analizirali fosil sa
intenzivnim laserom X-zrake iz Evropskog sinhroton radijacijskog
postrojenja na rubu Alpa. Proces je nazvan komputirana laminografija, u
kojem je 3D slika izrađena iz stotine 2D X-zrake-slika koje seku kroz
stvorenje. Tako da stvorenje može biti analizirano u finim detaljima bez
da ga se uništi.
Istraživači su napravili razumnu tvrdnju
da su pronašli femur (kost kuka), tibiu i fibulu (kost donje noge),
zglob kolena i kost članka. Dr. Houssaye je komentirala:
„Mi smo bili sigurni da je imao dve noge,
ali ih je bilo odlično videti i nadamo se da ćemo pronaći druge
karakteristike koje nismo mogli videti na drugom udu.“
Zaista, samo ime znači „zmija sa dobrim
nogama“, tako da čak i noge ustvari nisu bile novo otkriće; to je bio
detalj otkriven naprednom analitičkom tehnikom. Ali, da li „noge“
dugačke 2 cm i nedostajući prsti mogu biti klasificirani kao „dobro“. To
je drugo pitanje.
Dokaz evolucije?
Čak i sa pretpostavkom da može biti
utvrđeno da je predak današnjih zmija imao noge, kreacionisti u principu
nemaju problema sa gubitkom karakteristika kroz prirodne procese.
Razvoj beznožnosti nije dokaz evolucije molekula ka čoveku, koja zahteva
dodatak nove genetske informacije. Gubitak nogu može biti postignut
kroz degeneraciju DNK informacijske sekvence koja određuje razvoj nogu.
Vidi takođe: Beetle bloopers: Even a defect can be an advantage
sometimes.
Postoje dve protivničke evolucione
teorije. Jedna kaže da su se zmije pojavile od morskih reptila nazvanih
mosasauri, dok druga tvrdi da su se pojavile od kopnenih kopača tj.
monitor guštera. Istraživači se nadaju da će ovaj novi fosil razrešiti
debatu. Ali, ovo znači da karakteristike koje navodno pokazuju
zajedničkog pretka prema jednoj teoriji, moraju zapravo biti
homoplazije, tj. konvergentna evolucija karatkeristika koja se pojavila
nezavisno, ako je druga teorija tačna. Ali, homologija se smatra da je
dokaz za evoluciju (bez obzira na mnoge probleme; vidi: Common
structures = common ancestry?). Pozivanje na homoplaziju je zapravo
ublažavanje dokaza koji se ne uklapaju u paradigmu i zaista je takvo
ublažavanje dokaza sveprisutno.
Bolje objašnjenje je da su mosasaur
zastupnici u pravu da su zmije mogle evoluirati od monitor guštera i
zastupnici monitora su u pravu da su zmije mogle evoluirati od
mosasaura. To jest, zmije nisu evoluirale ni od čega i stvorene su kao
zmije!

Slika: news.bbc.co.uk
Kako je Eupodophis descouensi mogao izgledati. Noge su skroz dole na telu.
Kredibilna evoluciona priča treba da
pokazuje „primitivne“ zmije sa istaknutim nogama i dok su zmije
postajale „naprednije“ noge su se trebale smanjivati. Ali, fosilni zapis
zmija ne pokazuje to. Zmije koje su datirane kao „starije“ od nogatih
ne pokazuju da su imale noge. To jest, nogate zmije izgledaju kao
„napredne“ na određeni način. Prije opisana nogata zmija Haasiophis
terrasanctus je bila dovoljno „napredna“ da otvori vilicu da pojede plen
veći od njene glave isto kao što kopneni konstruktori kao pitoni i
danas rade. Ova i druge nogate zmije Pachyrhachis problematicus su bile
„napredne zmije tako da su ponovo evoluirale noge“, prema Olivier Repelu
iz Fild muzeja u Čikagu.4 Drugim rečima, on ne veruje da je ova jedinka
evolucioni prethodnik prve beznoge zmije.
Raž i Eskvil kažu za sve tri nogate zmije Haasiophis, Pachyrhachis i Eupodophis:
„tri zadnjonožne zmije imaju
makrostomatnu lobanju, ali u današnjim zmijama ova osobina se pojavljuje
samo u oblicima za koje se smatra da su „najnapredniji“, Makrostomata, a
priori, ova struktura treba biti izvedena.“5
Značajno da sve tri zmije su u jednoj stratografskoj fazi i jednoj geografskoj lokaciji. Raž i Eskvil kažu:
„Činjenica da potvrđeno ili izvedeno sve
nogate zmije su iz Cenomaijskog doba je iznenađujuće. … Dodatno, ove
zmije imaju veoma ograničenu geografsku distribuciju. Sve se pojavljuju u
„mediteranskom“ području Tetisa ili u njegovoj okolici, na severnim,
istočnim i južnim marginama postojećeg Mediterana i njegovom proširenju
sve do prelaznog područja između Akvitanije i Pariškog korita.“
Ovo podseća na posebnost da je jedna
provincija Kine izbacila sve navodne pernate dinosaure kao i Arheoraptor
prevaru, osim što istraživanju nogate zmije nedostaju upitni aspekti
pernate dino industrije.
Noge su sićušne u usporedbi sa veličinom
stvorenja za sve tri nogate zmije. Ali, čak i danas boa i pitoni imaju
sićušne „krakove“ koji se nalaze blizu njihovih krajeva koji se danas
koriste za hvatanje tokom parenja. Vidi takođe: Vestigal Organs: What do
they prove?
Moguće biblijski saglasno objašnjenje
Jedna ideja nije moguća, a to je da je
ovo zmija iz doba Pada kada je Bog prokleo zmiju u Postanju da gmiže na
svom stomaku (Postanje 3:14). Nije, s obzirom da je ovo fosil,
verovatnije je formiran u Nojevom Potopu oko 1.600 godina nakon
Prokletstva. Zapazi takođe, svaki geološki red nije redosled doba, nego
redosled zatrpavanja od strane Potopa i njegov posledični efekat. Vidi:
Biblical Geology: Properly Understanding the Rocks.
Ali, takvi fosili mogu osvetliti prirodu
Prokletstva. Mi ne znamo tačno šta je Bog učinio sa zmijom, ali jedna od
mogućnosti je da je ugasio genetsku informaciju koja čini noge, kukove i
druge karakteristike nužnim za hod.
Ako je ovo tako, mutacija je mogla
upaliti nešto od ove informacije ponovo, ali nepotpuno. Takođe,
nekorištene sekcije genoma će verovatnije akumulirati mutacije bez
prirodne selekcije da ih iskoreni. Tako da ako se ova sekcija upalila
pre hiljadu godina nakon Pada, mogla je biti upropašćena sa svim
mutacijama. Tako da je jedini rezultat u sitnim deformisanim nogama
previše malim za hod. Činjenica da su ove nogate zmije zatrpane u jednom
području i na istoj fazi u Potopu upućuje na zajednički okolišni faktor
koji je uticao na ovu ekološku zonu.
Dandanas, još više vremena od stvaranja,
čak i više mutacijskog tereta je moglo biti akumulirano značeći da bi
bilo čak i manje razumljive informacije da se upali. Dr. Džon Sendfor,
izumitelj genetskog pištolja je pokazao da stopa akumuliranog
mutacijskog oštećenja je tako velika da bi uništila naše genome u
potpunosti u navodnim milionima godina. Pogledaj njegovu knjigu: Genetic
Entropy and the Mystery of the Genome.
Zaista, isključivanje informacije se
dešava stalno, svaki put kad embrion uzrasta u odraslu jedinsku. Svaki
pojedinac započinje kao jedna ćelija – zigota ili kao ovum oplođen od
spermatozoida. Ovaj oplođeni ovum ima sve instrukcije kodirane u DNK da
nas napravi u ono što smo mi fizički (pod odgovarajućim okolišnim
uslovima).
Ali, kako embrion raste različite ćelije
na različitim mestima se moraju specijalizirati, tako da jedino određene
instrukcije su izvršene – ćelije postaju diferencirane. Instrukcije su
tamo, ali ugašene nekako, i u pravoj sekvenci. Prisutni su komplikovani
genetski prekidači i takođe proces nazvan metilacija – vezivanje metil
grupa hemijskim „slovima“ DNK koja kodiraju za instrukcije koje trebaju
biti „ugašene“.
Jedva da je protezanje predložti da Onaj
koji je programirao sekvence da se informacije gase tokom razvoja mogao
je i ugasiti informaciju u zmiji. Slično, jedno objašnjenje mesožderstva
koje se pojavilo nakon Pada je paljenje prikrivene genetske informaciju
za strukture odbranu-napad – vidi specifični primer žarećeg mehanizma
meduze.
Sažetak
Nekoliko zmija sa sićušnim nogama su otkrivene, ali svuda jedinstvene geološke lokacije i „doba“.
Istraživači su koristili naprednu tehniku da pokažu fini detalj u nogama jednog od njih, Eupodophis.
Gubitak strukture je konzistentan sa Padom, tako da nije dokaz evolucije nad stvaranjem.
Postoje dve nespojive teorije evolucije zmije, od vodenih reptila nazvanih mosasauri i od kopačkih guštera.
Noge postoje na zmijama koje su jedva
„primitivne“ čak i po evolucionom mišljenju. Tačnije, one imaju
karakteristike koje postoje samo kod zmija koje evolucionisti smatraju
„naprednim“. One se takođe pojavljuju „poznije“ u fosilnom zapisu nego
beznoge zmije. Neki evolucioni autoriteti njih ne smatraju da su preci
prvih beznogih zmija.
Kada je Bog prokleo zmije da hodaju na
trbuhu, On je verovatno ugasio genetsku informaciju za noge i druge
strukture. Nogate zmije mogu biti rezultat mutacije koja pali ovu
informaciju ponovo, ali ova informacija je bila oštećena mutacijama još
od Pada.
- Amos, J., Ancient serpent shows its leg, BBC News, April 2008.
- Rage J.C. & Escuillié F., Un nouveau serpent bipède du Cénomanien Crétacé. Implications phylétiques, Comptes rendus des’séances de l’Académie des Sciences, Paris, Series IIa, 330(7):513–520, 15 April 2000. Rage J.C. & Escuillié F. Eupodophis, 2002. A new name for the genus Podophis
- Rage and Escuillié, 2000, an extinct bipedal snake, preoccupied by Podophis Wiegmann, 1843 (Lacertilia, Scincidae), Amphibia-Reptilia, Leyden, vol. 23, pp. 232–233.
- Hecht, J., Prehistoric pins, New Scientist 2231:12, 25 March 2000. Return to text.
- Rage J.C. & Escuillié F., The Cenomanian: stage of hindlimbed snakes, Carnets de Géologie / Notebooks on Geology: Article 2003/01 (CG2003_A01_JCR-FE) .
Da li je Postanje poreklom od Enuma Eliš
str. 62 i 63, the Genesis account, teološki, istorijski i naučni komentar na Postanje 1-11,
Džonatan Sarfati, PHD, F.M.
Najomiljenija teorija poticanja od pagana je da je Postanje posuđeno iz Enuma Eliša-a. To je mit napisan akadskim klinastim pismom koji se sastoji od 1000 stihova i zapisan je na sedam glinenih pločica. Ime dolazi od prvih reči „kada na visini“ – enuma eliš la nabu šamani, „kada nebesa iznad ne behu nazvana …“
Engleski istraživač i arheolog Ostin
Henri Lajar (1817-1894) otkrio je njegove fragmente u Ninevi (danas u
modernom Iraku). Engleski asirolog Džorž Smit (1840-1876), koi je takođe
otkrio Gilgamešov ep objavio ih je 1876. godine pretenciozno kao
Haldejski izveštaj Postanja.
Nakon što je ovo otkriveno Fridrih Delič
(1850-1922) je predložio tvrdnju da je većina Postanja posuđeno iz ove
knjige. Njegov prvi predlog je bio na predavanju Deutsche Orient
Gesellschaft u Berlinu pod nazivom „Babel und Bibel“ („Vavilon i
Biblija“), 13. januara 1802. godine. Fridrih Delič je bio sin Franca
Deliča koji je bio ko-autor odličnog komentara kojeg sam često citirao.
Nažalost, Fridrih je bio liberalan teolog nasuprot njegovom
konzervativnom ocu i takođe je napustio očev istaknut pro-semitizam.
Njegov učenik Paul Haupt (1858-1926) je čak bio zagovornik proto-naci
viđenja da je Isus arijskog porekla.[1]
Enuma Eliš nije ustvari uopšte izveštaj
stvaranja. On započinje sa pitkom vodom koja je oličena kao bog Apsu,
mešajući se sa slanom vodom oličenom kao boginja Tiamat. Upravo kad se
pitka voda uliva u more mulj se često stvara baš u delti i može biti to
tumačeno kao događaj stvaranja. To je zapravo jednostavna fizička
hemija: čestice mulja su već bile prisutne tamo u rečnoj vodi, ali
zahvaljujući nošenjem jednakih naelektrisanja, ove čestice su
suspendovane elektorostatskom repulzijom među njima. Kada se reka
susreće sa slanom vodom joni uveliko neutralizuju naelektrisanje i
poništavaju električnu repulziju. Tako da čestice flokuliraju (ugrubo:
talože se).[2]
Ali mit kaže da su bogovi rođeni od
sjedinjenjem Apsu i Tiamata. Međutim, kasnije oni čine tako veliku buku
da Apsu poželi da ih pobije. Najmoćiniji bog Ea uspeva da uspava Apsua i
potom ga ubija. Ali zatim Tiamat želi osvetu za ubistvo njenog muža i
rađa 11 bogova-čudovišta. Jedan, Kingu, postaje novi suprug Tiamata i
general njene armije.
Bogovi su preplašeni, ali tada Marduk sin
od boga Ea koji je čak i snažniji od Ea prihvaća da pomogne u zamenu za
to da bude proglašen Kraljem bogova. On hvata ogromnu Tiamat u mrežu,
usiljava žestoki vetar u njena usta koji naduvava njenu unutrašnjost i
ubija je sa strelom kroz njen stomak i srce. Tada razbija njenu glavu sa
toljagom, raseca njeno telo na dvoje „kao ribu za sušenje“ i pravi nebo
od gornjeg dela i zemlju od donjeg tela. Tiamatove plačne oči postaju
izvor reka Tigrisa i Eufrata i rep postaje Mlečni put.
Tada Marduk hvata Kingua i pogubljuje ga.
Od njegove krvi pomešane sa glinom stvoreni su ljudi da služe bogovima.
Marduk uz to postaje glavni bog Vavilona. U Hebrejskoj Bibliji je
poznat kao Merodah.
Kao što vidimo ovi „bogovi“ su smrtni,
nasilni i krhki, i nisu vrhovni gospodari nad tvorevinom. Ovo nije ništa
kao Tvorac Bog Postanja, vrhovni gospodar Bog nad svim što je stvorio.[3]
[1] Haupt, P., The Aryan Ancestry of Jesus, The Open Court 4(1):193-204; opensiuc.lib.siu.edu/ocj/vol1909/iss4/1
[2]
Ovo je jako pojednostavljen opsi onoga što je rigorozno objašnjeno
poznate DLVO (Deryagin-Landau/Verwey-Overbeek) teorije koloidne
stabilnosti.
[3]
Vidi takođe Adamthwaite, M.R., Is Genesis 1 just reworked Babylonian
myth? J.Creation 27(2):99-104, 2013. Murray Damthwaite, doktor sa
Melburn Univerziteta (Australia) u Drevnoj istoriji, jeszicima
(akadskim) i književnosti.
Mojsije - autor Petoknjižja
str. 9 i 10, the Genesis account, teološki, istorijski i naučni komentar na Postanje 1-11
Džonatan Sarfati, PHD, F.M.
Prvih pet knjiga Biblije – Postanje,
Izlazka, Levitska, Brojevi i Ponovljeni zakoni – su poznate kao Tora,
Zakon ili Petoknjižje. Tora je odavno bila pripisana Mojsiju. Zaista,
unutrašnji dokaz iz knjiga ukazuje na Mojsija, jer one tvrde da je on
autor, npr. Izlazka 17:4; 24:4-7; 34:27; Brojevi 33:2; Ponovljeni zakoni
31:9, 22, 24.
Na ovoj potpori, druge knjige Staroga
Zaveta potvrđuju da je Mojsije autor, npr.Isus Navin 1:7-8; 8:32-34;
Sudije 3:4; 1 Careva 2:3; 2 Careva 14:6; 21:8; 2 Hronike 25:4; Jezdra
6:18; Nemija 8:1; 13:1; Daniel 9:11-13.
Nastavljajući u Novi Zavet, njegovi pisci
takođe potvrđuju da je Mojsje autor, npr. Jovan 1:17; Dela Apostolska
6:14; 13:39; 15:5; 1 Korinćanima 9:9; 2 Korinćanima 3:15: Jevrejima
10:28.
Konačno, sam Isus navodi Mojsije kao
autora. On često govori o Mojsijevim spisima ili Zakonu Mojsijevom, bez
ikakvog poricanja, npr. Matej 8:4; 19:7-8; Marko 7:10; 12:26; Luka
24:27, 44; Jovan 7:19. Zaista, Isus naglašava ozbiljnost odricanja
Mojsija nekoliko puta.
Isus je upozorio neverujuće Jevrejske vođe u Njegovo vreme:
„Jer da verovaste Mojsiju, verovali biste i meni; jer on pisa o meni.
A kad njegovim Pismima ne verujete, kako ćete verovati mojim rečima?“
Jovan 5:46-47
Slično i danas, liberalni teolozi koji
sumnjaju Mojsiju često sumnjaju i šta je Isus rekao (osim ako selektivno
korištenje Njegovih reči ne može nekako biti izvrnuto da daje potporu
političko korektnom cilju s kojim se oni slažu). Čak još važnije, u
izveštaju o bogatom čoveku nakon smrti u Luka 16:31, Isus kazuje kako je
ovaj čovek molio nekoga da se vrati iz mrtvih da upozori njegovu braću.
Ali kako Isus objašnjava dalje:
"On (Avram) mu je rekao (bogatom čoveku u
Paklu),“Ako ne slušaju Mojsija i proroke, ako neko i iz mrtvih vaskrsne,
neće se uveriti.“
Ovo je izuzetno ozbiljno: Isus je rekao
da oni koji odbace Mojsija neće biti uvereni čak i samim vaskrsenjem.
Tako da nije čudno da one crkve i seminari koji odbacuju istoričnost
Mojsijevih spisa često takođe odbacuju doslovno telesno vaskrsenje
Gospoda Isusa Hrista.
Zaista, Isus je otišao dalje u Matej 5:18 da potvrdi apsolutni autoritet Mojsijevih spisa, Zakona:
„Jer zaista vam kažem: Dok ne prođe nebo i
zemlja, neće nestati ni najmanjega slovca ili jedne crte iz Zakona dok
se sve ne zbude.“[1]
Isus potvrđuje inspiraciju Pisma čak i do
najmanjeg slova – grčke jote, ekvivalenta hebrejskom jod („jot“ KJV) –
ili dela „crte“, najmanjeg poteza pisaljke odnoseći se na beta, ili
kapa, ili daleta, ili reša („naslova“ – KJV).
[1]
Primenjivost Mojsijeovg Zakona za danas je drugo pitanje od njegove
nepogrešivosti i njegovog autoriteta njegovoj namenjenoj publici, i
izvan područja CMI. Ali vidi Cosner, L., Is eating shellfish still an
abomination? creation.com/all-food-clean, 1. septembar 2012.
Dokaz Boga – kozmološki argument
Postoje razni dokazi Boga, kako
filozofski, tako i naučni i istorijski. Ovde ću se osvrnuti na
filozofski dokaz Boga i to jedan od njih tzv. kozmološki dokaz Boga ili
jedan od njih.
Kozmološki dokaz dokazuje Boga na
principima stvaranja. Kao prvo da bi se dokazalo da vreme ne ide unazad u
beskonačnost možemo se poslužiti Kalam argumentom. Ako uzmemo niz
domina koje su poredane jedna kraj druge i one padaju u niz i ako kažemo
da je današnje vreme vreme kada je određeni niz domina pao, ako idemo
dalje u prošlost mora biti vreme kad su domine započele da padaju. Znači
da bi došli do početka vremena dolazima do početka padanja domina.
Stoga je nužno da su domine jednom počele da padaju da bi domine došle
do ovog vremena u kojem se nalazimo. Znači postoji početak vremena. Onda
na osnovi tog agumenta dolazimo do početka vremena kada ničega nema. A
kako je sve nastalo što danas postoji? Ništa ne može nastati samo od
sebe. Stoga da bi svemir nastao mora biti stvoren, jer nešto ga mora
stvoriti. Početak materije mora biti nematerijalan i bezgranično moćan.
Početak celog kosmosa mora biti kompleksniji od samog kosmosa. Samo
komleksnije stvara manje kompleksno. Znači, tvorac svega je Bog. Početak
vremena je večan. Tvorac je bezgranično moćan.
Na koji način kompleksnije stvara manje
kompleksno. Evo primera, da bi nastalo dete trebaju oba roditelja, da bi
nastalo umetničko delo mora ga oslikati umetnik koji je kompleksniji od
samog umetničkog dela.
Napad na kozmološki argument je napravio
Kant tvrdeći da se iz empritijskog, iskustvenog prelazi u transcadentno.
Međutim, to nije pobijanje dokaza već samo opis argumenta. Sasvim je
irelevantno što se empirijski dokazuje trancedentno, jer se putem
argumenta samo dokazuje postojanje transcadentnog tj. Boga, a ne ulazi u
teološke opise i dubine, ovde se samo dotičemo trancadentnog, a ne
ulazimo u teološko dokazivanje. Ne idemo u teološko opisivanje, u
dubine. Sasvim je nebitno što se iz empirijskog prelazi u trancadentno,
jer se putem empirijskog ovde dokazuje trancadentno, ali se ne ulazi u
dubinu. Ako ja npr. mogu dokazati trancadentno putem empirijskog sasvim
je irelevantno što se to obavlja, dakle, to je samo opis argumenta, a ne
njegovo pobijanje. Ako npr. neko može dokazati putem dokaza ko je ubio
žrtvu, sasvim je irelevantno što ti pomoću dokaza možeš da pronađeš
ubicu koga nikad nisi ni video, niti ga znaš, znači sasvim je nebitno
što inspektor ne vidi ubicu niti ga zna, a može da dokaže da je neki
čovek ubica. Kao i na primer kad bi čovek mogao da hoda i da leti,
sasvim je nebitno što sad čovek hoda, pa sad opet leti, i ako neko kaže:
„on od hodanja prelazi na letenje, znači to nije istinito“; to je
besmislica, jer ako neko to radi tj. prelazi od hodanja na letenje
nikakav značaj nema ta kritika ili takoreći pobijanje, jer on to čini.
Stoga, to je samo opis argumenta, a ne njegovo pobijanje. Ako neko može
kozmološkim argumentom dokazati Boga, sasvim je nebitno što on putem
iskustvenog dokazuje trancadentno. Tako da Kant nije u pravu.
Kozmološki argument stoji, a Kant se pokazuje sofistom.
Neuspeh radioaktivnog „datiranja“
Nedavni isticanje lave u Novom Zelandu pokazuje „starost“ od miliona godina
Endrju Sneling
Članak iz Creation 22(1):18-21-decembar 1999.
Nalazeći se otprilike u centru
Severnog ostrva u Novom Zelandu, Mt Ngauruhoe je najnoviji vulkan i
jedan od najaktivnijih na Novom Zelandu (Slika 1 i 2). Nije tako dobro
publiciran kao njegov veći bliži sused Mt Ruapehu koji je eruptirao
otprilike nekoliko puta u poslednjih pet godina.
Međutim, Mt Ngauruhoe je upečatljiv,
gotovo savršena kupa koja se uzdiže više od 1.000 metara (3.300 stopa)
iznad okružujućeg pejzaža do uzvišenja od 2.291 m (7.500 stopa)
nadmorske visine1 (Slika 3). Erupcije iz centralnog 400 m
(1.300 stopa) širokog kratera su stvorile strmost kupole (od 33 stepeni)
vanjskog nagiba.

Slika 1. Lokacija Mt Ngauruhoe, središte Severnog ostrva, Novi Zeland
Mt Ngauruhoe se smatra da je bio aktivan
barem 2.500 godina, sa više od 70 erupcijskih perioda od 1839. godine
kada su evropski doseljenici prvi put zabeležili erupciju pare.2
Naravno, prije toga, Maori su prisustvovali mnogim erupcijama iz
planine. Prva erupcija lave viđena od Evropljana se desila 1870. godine.3
Sledeće su bile erupcije pepela svakih nekoliko godina sve do glavne,
eksplozivne erupcije u aprilu-maju 1948. godine, nakon čega je usledilo
isticanje lave niz severnozapadnu strminu februara 1949. godine.2,3 Procenjena zapremina lave je bila otprilike 575.000 m3 (20 miliona kubnih stopa).
Erupcija koja je trajala od 13. maja
1954. godine do 10. marta 1955. godine je započela eksplozivnim
izbacivanjem pepela i blokova.2,3 Nakon toga gotovo 8 miliona m3 (280 miliona kubnih stopa) lave je isteklo iz kratera u serijama od 17 različitih isticanja sledećih datuma 1954. godine:
– Jun 4, 30
– Jul 8,9,10, 11, 13, 14, 23, 28, 29, 30
– Avgust 15(?), 18
– Septembar 16, 18, 26
Fotografija Krejg Potona

Slika 2. Pogled iz ptičje perspektive, gledajući južno od izlasku sunca, vulkani Mt Ngauruhoe (ispred) i Mt Ruapehu (u pozadini).
Ova isticanja su još i danas vidljiva na
severozapadnoj i zapadnoj strmini Ngauruhoe (Slika 4). Isticanje 18.
avgusta je bilo više od 18 m (55 stopa) debelo i još toplo gotovo nakon
godinu dana od očvrsnuća. Eksplozije pepela su završile ovaj dugi
erupcijski period.
Nakon toga, Ngauruhoe je izbacivao paru gotovo stalno, sa mnogim malim eupcijama pepela2
(Slika 5). Topovski slične erupcije u januaru i martu 1974. godine su
izbacile napolju veliku količinu pepela, kao stub u atmosferu i to dok
je odron tekao dole sa strana kupole. Blokovi koji su teški do 1.000
tona su izbačeni 100 m (300 stopa). Međutim, najsnažnija eksplozija se
desila 19. februara 1975. godine, praćena onim što su očevidci opisali
kao atmosferske udarne talase.4 Blokovi do 30 m (100 stopa)
širine su katapultirani do 3 km (skoro 2 milje). Erupcijsko pero je bilo
11-13 km (7-8 milja) visoko.
Fotografija Endjru Snelinga

Slika 3. Mt. Ngauruhoe viđen severno od Mt. Ruapehu.
Turbulentne lavine pepela i blokova su zapljuskivale strane Ngauruhoea od oko 60 km (35 milja) po satu.2 Procenjeno je da je barem 3,4 miliona m3 (120 miliona kubnih stopa) pepela i blokova izbačeno u 7 sati.4 Od tada nisu se desile naknadne erupcije.
Datiranje stena
Radioatkivno datiranje, uopšteno zavisi od tri glavne pretpostavke:
- Kada se stena formira (očvrsne) trebaju biti prisutni u steni samo radioaktivni atomi roditelja i nimalo radiogenskih (koji nastaje radioaktivnim raspadom drugog elementa) atoma kćerke;5
- Nakon očvsnuća, stena mora ostati zatvoreni sistem, to jest, nimalo roditelja ili atoma kćerke ne smeju biti dodani ili oduzeti iz stene vanjskim uticajem kao npr. prodiruća podzemna voda; i
- Brzina radioaktivnog raspada mora ostati konstantna.
Ako je bilo koja od ovih pretpostavki prekršena, tada tehnika propada i bilo koji rezultati „datiranja“ su pogrešni.
Fotografija Endjru Snelinga

Slika 4. Pogled iz
Mangateopopo doline na podnožju Mt Ngauruhoe, pokazuje tamnije-obojano
nedavno isticanje lave na severozapadnim strminama.
Kalium-argon (K-Ar) metoda datiranja se
često koristi za datiranje vulkanskih stena (i kao dodatak, obližnjih
fosila). Korištenjem ove metode, pretpostavlja se da nije bilo kćerke-
radiogenski argon (40Arx) u steni kada je stena formirana.6
Za vulkanske stene koje se hlade od tekuće lave, ovo bi izgledalo kao
razumna pretpostavka. S obzirom da je argon gas, on bi trebao pobeći u
atmosferu zbog intenzivne toplote lave. Naravno, nijedan geolog nije bio
prisutan da testira ovu pretpostavku promatranjem drevne lave kada se
hladila, ali mi možemo proučavati sadašnja isticanja lave.
Kalium-argon „datiranje“
Jedanaest uzoraka su prikupljeni od pet
nedavnih isticanja lave tokom rada na terenu januara 1996.-po dva od
isticanja od 11. februara 1949. g., 4. juna 1954. g. i 14. jula 1954. g.
i od 19. februara 1975. godine7 (Slika 6). Mračnije nedavne
lave su bile jasno vidljive i svaka je lako identifikovana (s pomoću
mapa) na severnozapadnim strminama nasuprot svetlije-obojanim delovima
kupe (Slike 4 i 7). Sva isticanja su tipično pretvorena u nabacane
blokove stvrdnute lave, rezultirajući u grube, reckave, ugljenisane
površine (Slika 8).
Fotografija Džima Hilija

Slika 5. Mala erupcija pepela, Mt Ngauruhoe.
Slika 6. Dodatak:
Andezit isticanja 30. juna 1954. g., Mt Ngauruhoe viđen 60 puta povećan
pod geološkim mikroskopom. Različiti minerali imaju različite boje. Svi
su utisnuti u mrežu fine prašine.
Uzorci su poslani nizom u serijama
Geohron labaratoriji u Kembridžu, Boston (SAD), za celokupno datiranje
stene kalium-argon (K-Ar) datiranja – prvo deo po jednog uzorka od
svakog isticanja, zatim deo drugog uzorka iz svakog isticanja nakon
primitka rezultata prvog seta i konačno deo trećeg uzorka od isticanja
30. juna 1954.7 Da bi takođe testirali konzistentnost
rezultata iz uzoraka, drugi delovi od dva uzorka lave od 30 juna 1954.
godine su takođe poslani na analizu.
Geohron je ugledna komercijalna
labaratorija, K-Ar labaratorijski direktor ima doktorat u K-Ar
datiranju. Nijedna posebna lokacija ili očekivana starost nije navedena
labaratoriji. Međutim, uzorci su opisani kao moguće mladi sa veoma malo
argona u njima tako da bi osigurali da je posebna briga vršena tokom
analitičkog rada.
„Datiranja“ pribavljena iz K-Ar analiza
su uvršteni u Tabliccu 1.7. „Starosti“ dosežu od <0,27 do 3,5 (±0,2)
miliona godina za stene koje su promatrane da su se ohladile od lave pre
25-50 godina ranije. Jedan uzorak od svakog isticanja lave je dao
„starost“ od <0,27 do <0,29 miliona godina dok su svi ostali
uzorci dali „starost“ u milionima godina. Uzorci male „starost“ su svi
procesuirani od labaratorije u istoj seriji, upućujući na sistematski
labaratorijski problem. Tako da je direktor labaratorije ljubazno ponovo
proverio svoju opremu i ponovo obavio testiranje uzoraka, dajući
slične rezultate. Ovo je objasnilo sistematsku labaratorijsku grešku i
potvrdilo da su niski rezultati ispravni. Dalje, ponovljena merenja na
uzorcima već analiziranim (A#2 i B#/2 u Tablici 1) nisu pokazali iste
rezultate, ali ovo nije bilo iznenađujuće objašnjavajući analitičku
nesigurnost na tako niskom nivou argona. Očito, sadržaj argona varira
uveliko unutar ovih stena. Neki geohronolozi bi rekli da je <0,27
miliona godina zapravo ispravan „podatak“, ali kako bi oni znali da
ustvari 3,5 miliona godina nije ustvari prava „starost“ ako nisu već
znali da je tečenje lave bilo nedavno?!
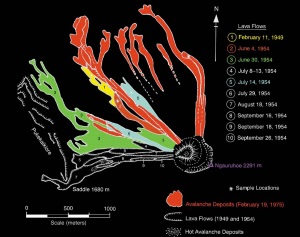
Slika 7. Mapa sevenozapadne strmine Mt Ngaruhoe koja pokazuje tečenje lave 1949. g. i 1954. i 1975. g. nakupine odrona.3,4
Zato jer su ove stene poznate da su mlađe
od 50 godina, očito je iz analitičkih podataka da ove K-Ar „starosti“
su zbog „viška“ argona koji je nasleđen iz područja magma izvora duboko u
zemlji.7 Stoga, kada se lava ohladila ona je sadržavala prihvatljivu (ne nula) koncentraciju „normalnog“ 40Ar, koji se ne razlikuje od kćerke – radiogenskog 40Arx koji je dobijen raspadanjem radioaktivnog roditelja – 40K.
Ovo krši pretpostavku (1) radioaktivnog raspada, i tako K-Ar metod pada
na testu. Isti neuspeh je rakođe poznat da se dešava u mnogim drugim
stenama, uključujući ujedno nedavno vulkanske8 i drevne koraste stene.9
Fotografija Endrju Snelinga

Slika 8. Isticanje lave od 30. jun 1954., pokazuje razbacane blokove formirane lave, koja daje grubu, reckavu i ugljenisanu površinu.
Zaključak
Metoda radioaktivnog datiranja kalium-argon se pokazala neuspešnom na isticanja lave na Mt Ngauruhoe, Novi
Zeland od 1949., 1954. i 1975. godine, bez obzira na kvalitet
labaratorijskog K-Ar analitičkog rada. Gas argon koji je iznesen iz
dubine unutar zemlje, unutar istopljene stene je već bio prisutan u
lavama kada se hladila. Mi znamo pravu starost stena, jer su one
promatrane da su se formirale prije manje od 50 godina. Međutim, one
pokazuju „starosti“ do 3,5 miliona godina koje su stoga pogrešne. Kako
se mi možemo pouzdati korištenju ovoga istog metoda „datiranja“ na
stenama čiju starost ni ne znamo? Ako je metod neuspešan na stenama kada
imamo verodostojan izveštaj očevidaca, onda zašto bi imali pouzdanje u
taj metod za druge stene gde nema nijedne verodostojne istorijske
provere?
Međutim, mi znamo nekoga koje je bio
prisutan kada su se sve zemljine stene formirale-samog Tvorca. On nam je
rekao kada je to bilo u svom izveštaju očevidca u prvoj knjizi Biblije,
Postanju, tako da znamo koliko su stene stare. Koliko bolje da stavimo
svoje pouzdanje u Tvorca koji je stvorio i zna sve i koji nikada ne
greši ili govori laži nego u radioaktivnu metodu datiranja koja je
ponovljeno pokazivana kao pogrešna i da pokazuje pogrešnu starost za
zemljine stene.
K-Ar (kalium-argon) metod datiranja
Fosili se gotovo nikada ne datiraju
radiometričnim metodama s obzirom da oni retko sadrže odgovarajuće
radioaktivne elemente. Uobičajeni način datiranja fosila (i stena koje
ne sadrže radioaktivne elemente) je putem „datiranja“ povezane vulkanske
stene. Ovo je uobičajeno rađeno koristeći K-Ar metod. Ono ovisi o brzini
po kojoj radioaktivni kalium se raspada u gas argon.
K-Ar metod radi na pretpostavci da „sat“
počinje da „kuca“ momentom koji se stena očvrsne. To jest, pretpostavlja
se da nimalo argona dobijenog radioaktivnim raspadom nije prvobitno
bilo prisutno, ali tek nakon što se lava ohladila i očvrsla argon iz
radioaktivnog raspada nije mogao da pobegne i započeo je da se
akumulira. Međutim, poznato je da ako se radioaktivno „datiranje“
suprotstavlja (evolucionoj) starosti izvedenoj iz fosila, podatak se
odbacuje kao pogrešan. Vidi Lubenov, M, The Pigs took it all, Creation
17(3):36–38, 1995.
| Datum isticanja | Uzorak | Labaratorijski kod | K-Ar „starost“ (milioni godina) |
| 11 februar 1949. | A | R-11714 | < 0,27 |
| B | R-11511 | 1,0 ± 0,2 | |
| 4 jun 1954. | A | R-11715 | <0,27 |
| B | R-11512 | 1,5 ± 0,1 | |
| 30 jun 1954. | A #1 | R-11718 | <0,27 |
| A #2 | R-12106 | 1,3 ± 0,3 | |
| B #1 | R-12003 | 3,5 ± 0,2 | |
| B #2 | R-12107 | 0,8 ± 0,2 | |
| C | R-11513 | 1,2 ± 0,2 | |
| 14 jul 1954. | A | R-11509 | 1,0 ± 0,2 |
| B | R-11716 | <0,29 | |
| 19 februar 1975. | A | R-11510 | 1,0 ± 0,2 |
| B | R-11717 | <0,27 |
Tablica 1. Kalium-argon „datiranja“ nedavnih Mt Ngauruhoe (Novi Zeland) isticanja lave.7
Beleška ed. Ovaj članak
iz Creation magazina od dr. Snelinga je zasnovan na njegovom tehničkom
radu, Ref 7, koji je puno više detaljniji o metodi istraživanja i
odgovara mogućim kritiziranjima koje je moglo biti u Creation magazinu.
Reference i beleške:
- Williams, K., Volcanoes of the South Wind: A Field Guide to the Volcanoes and Landscape of the Tongariro National Park, Tongariro Natural History Society, Turangi, New Zealand, 1994.
- Nairn, I.A. and Wood, C.P., Active Volcanoes and Geothermal Systems, Taupo Volcanic Zone, New Zealand Geological Survey Record 22:5–84, 1987. Return to text.
- Gregg, D.R., The Geology of the Tongariro Subdivision, New Zealand Geological Survey Bulletin n.s.40, 1960.
- Nairn, IA and Self, S., Explosive eruptions and pyroclastic avalanches from Ngauruhoe in February 1975, Journal of Volcanology and Geothermal Research 3:39–60, 1978.
- Ovo je istina za K-Ar datiranje, jedne od najuobičajenijih metoda, i koja je ovde raspravljena. Takozvana "izohron" tehnika koja se bavi hemijskim analizama kamena koji je "datiran" nastoji da izbegne ovu prepostavku. Rasprava o izohrom "datiranju" kao i povezanim problemima lažnim (pseudo) izohronima, je van područja ovog naučnog rada, ali pogledaj Austin, S.A. (ed.), Grand Canyon: Monument to Catastrophe, Institute for Creation Research, Santee, California, pp. 111–131, 1994.
- Dalrymple, G.B., The Age of the Earth, Stanford University Press, Stanford, California, p. 91, 1991.
- Snelling, A.A., The cause of anomalous potassium-argon ‘ages’ for recent andesite flows at Mt Ngauruhoe, New Zealand, and the implications for potassium-argon ‘dating’, In: Walsh, R.E. (ed.), Proceedings of the Fourth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, Pennsylvania, pp. 503–525, 1998.
- Snelling, AA, Excess argon’: the ‘Archilles’ heel’ of potassium-argon and argon-argon ‘dating’ of volcanic rocks, Institute for Creation Research, Santee, California, Impact #307, 1999.
- Snelling, AA, Potassium-argon and argon-argon dating of crustal rocks and the problem of excess argon, Institute for Creation Research, Santee, California, Impact #309, 1999.
Baraminologija – klasifikacija stvorenih organizama
Autor: Vejn Frejr
CRSQ, vol 37, No 2, str. 82-91, septembar 2000.
Abstrakt
Decenijama su kreacionisti koristili reč
„rod“, „tip“ ili „grupa“ za svoje prihvaćene kategorije genetski
nepovezanih organizama koji uključuju sve one oblikovane od Tvorca tokom
nedelje stvaranja. Unutar svake od ove kategorije različite vrste,
podvrste i varijacije su smatrane da su se razdvojile od zajedničkog
pretka. Međutim, sve do prije nekoliko godina nije bilo ozbiljne opsežne
metodologije klasifikacije usredsređene na karakteriziranju svake
prvobitne kategorije koja je odvojena genetskim prazninama od svih
drugih kategorija. Sada se baraminologija (sa sistematikom
diskontinuiteta) razvila u plodan pristup u klasifikaciju unutar
kreacionističnog modela. Terminologija i metodologija se razvila i prva
naučna konferencija baraminologije je održana u leto 1999. godine.
Agresivan budući program je zamišljen.
Uvod
Osnovne ljudske karakteristike uključuju
klasifikaciju i identifikaciju. Ljudi rade to toliko često da je
činjenje toga suštinsko našem načinu života. Pojedinci su naučili
klasifikaciju misli i stvari i dok vrše interakciju sa okolinama oni
uporno identifikuju spoznaju kao miris, prizore (nebo, hrana, životinja,
biljka, itd.), zvukove (zvuk trube, lajanje psa, muzika, itd.) ukuse
(slatko, ljuto, itd.), i druge utiske kao grubo i glatko. Kompleksnost
svega ovoga je zapanjujuća.
Zanimljivo, Biblija uključuje izveštaj o
tome kako je Bog doveo Adamu sve kopnene životinje i ptice (Postanje
2:19) tako da ih Adam može promotriti i imenovati ih. Ova priča može da
ima dublji smisao nego što površno čitanje otkriva. U procesu imenovanja
stvorenja Adam je naučio nešto o sebi. On je shvatio svoj kapacitet da
shvati i otkrije šablon. Neka su stvorenja imala ljuske, a neka dlake
ili perije.
Dodatno, Adam je naučio nešto o Bogu –
Dizajneru. U stvaranju je bio izraz Božijeg stvaranja. Nakon što je Adam
otkrio prirodu on nikada neće biti isti kao prije. Zvezde, životinje,
biljke – sve su bile deo Božijeg stvaranja. Pozornica je postavljena i
posledično tome milijarde učenika nauke će igrati deo na pozornici
života i tokom tog vremena osećat će uzubuđenje otkrivanja Božije poruke
u prirodi (Psalam 19:1, Rimljanima 1:19-20; i vidi Remajn, 1993).
Taksonomija i sistematika
Promišljajući o ovim stvarima čini nam se
lakše da shvatimo osnovnu važnost klasifikacije u biološkim studijama
izumrlih i postojećih oblika života. Taksonomija je termin
korišten za nauku klasificiranja živih stvari shodno njihovim prirodnim
grupisanjima. Suštinski, naučnici danas koriste taksonomski sistem
predstavljen od strane švedskog botaniste Linea, pre 250 godina.
Međutim, postoje različiti pristupi korišteni od naučnika za proučavanje šablona života i za klasifikaciju. Oni se nazivaju sistemi taksonomije i oni leže pod opštim naslovom sistematike.
Za proučavanje šablona u prirodi Remajn (1993. str. 444) je usporedio
različite procedure sa funkcijama različitih tipova filma (infracrveni,
ultraljubičasti, i film x zrake). Na primer, postoje filetske
ili evolucione sistematike (ili Darvinske sistematike) u kojima se
makroevolucija (uključuje velike promene) pretpostavlja. Vidi sliku 1.
Sistematske šeme koje stavljaju manje naglaska na evoluciju mogu biti nazvane fenetske,
jer su zasnovane na karakteristikama organizama i ne nužno njihovim
mogućim velikim razmerama evolucionih odnosa. Takođe, postoji i kladistički
pristup koji se usredsređuje na zajedničkim ili tzv.
„deljenim/izvedenim“ osobinama. Najvažnije, popularna metodologija u
filetskim, fenetskim i kladističkim procedurama, sve su prilagođene makroevolucionom teoretisanju.
Najranije metode su bile fenetske, koje
su dale osobitu prednost bivajući objektivno opažljive i
najproverljivije. Kritičari osnovne fenetike osećaju da njegova
jednostavnost ne ohrabljuje adekvatno razmatranje istorijske
kauzalnosti. Bilo kako bilo, oni koji koriste baraminologiju (sa
sistematikom diskontinuiteta) kako je predstavljeno u ovom naučnom radu,
mogu biti agnostični u vezi popularne fenetične i kladistične
metodologije, ali oni barem uključuju neku fenetičnu metodologiju kao
jedan od njihovih alata.
Sistematika diskontinuiteta i barminologija
Naučnici koji su preferirali neki drugi, a
ne evolucioni model nedavno su razvili ono što oni veruju da je puno
realističnija sistematika zasnovana na nepovezanosti ili tipologiji
pronađenoj u prirodi. Metodologija je prikladno bila nazvana sistematika diskontinuiteta (ranije predstavljena od Valter Dž. Remajn, 1990.), ili kada je kombinovana sa biblijskim otkrivenjem baraminologija
(termin predstavljen od Kurt P. Vajza, 1990.). Baraminologija može biti
definisama kao taksonomija zasnovana na stvorenim rodovima (vidi Barc,
1991.; Frejr, 1991.; 1999.; i slika 2). Reč „barmin“ je smišljena od
Frenk L. Marša i prvo je objavljena 1941. godine, ona je izvedena od
hebrejskog glagola bara, stvoriti i min, rod (takođe vidi Marš, 1969.; Viliams, 1997.).
S obzirom da klasifikacija podvači sva
biološka istraživanja, prilično je značajno da kreacionisti sada imaju
aktivno usredsređenje na ovu temu. Suštinski napredak je napravljen od
1990. godine i baraminolozi su razvili svoju vlastitu terminologiju koja
se u ovo vreme čini da je prilično praktična za one koji se bave
sistematskim istraživanjima. Glavna svrha baraminologije je da odredi
koji organizmi dele zajedničko poreklo.
Marš je koristio termin baramin
na uključujući način za celokupnu grupu znanih, neznanih, i moguće
uključujućih organizama koji dele genetsku povezanost. Ali, sada je
fokus specifičniji i samo oni primerci koji mogu biti proučavani, isto
tako živi i izumrli (uključujući fosile) primerci mogu biti uključeni u
sadašnje četiri glavne baraminske grupe. Termini korišten kao četiri
primarne baraminske kategorije su: holobaramin, monobaramin, apobaramin, i polibaramin.
Holobaramini
U baraminologiji primarni termin je holobaramin od grčkog holos
za celo. Holobaramin su svi i samo oni poznati živi i/ili izumrli
oblici života shvaćeni da dele genetsku povezanost. To je celokupna
grupa za koju se veruje da je povezana zajedničkim poreklom.
Slika 1
Slika 2
Tako da sada svaka prirodna grupa
povezanih biljaka ili povezanih životinja čini holobaramin ili u još
specifičnijoj kreacionističkoj terminologiji holobaramin se sastoji od
svih poznatih organizama u grupi koji započinju nakon što je Bog stvorio
prvobitne organizme (vidi Vajz, 1991.). Holobaramin može biti
predstavljen kao granajuće drvo, stecišta i vrhovi grana predstavljaju
sve poznate članove (podvrste, vrste, itd.) „roda“ („grupe“ ili „tipa“).
Vidi sliku 2. Kada su jedinke ili grupe očito povezanih primeraka
uspoređeni mogu biti imenovani kao holobaraminski ako sačinjavaju delove jednog holobaramina.
Tokom nekoliko poslednjih decenija
kreacionistički istraživači su koristili termine „rod“, „grupa“ i „tip“
uopšte naizmenično ili su kao pojedini istraživači preferirali jedno ili
drugo osobito uopšteno ime za Maršov „baramin“ i takođe za šta se
osobitije i trenutno mogu biti imenovani kao holobaramini. Sada
sistematici određenog taksona biljkaka ili životinja mogu odbaciti
stariju terminologiju i napraviti njihova drva pokazujući holobaraminske
srodstvo i tako granice zajedničkog porekla.
Važan primer holobaramina bi bili ljudi,
Homo sapiens. Na vrhovima holobaraminskih grana su različite rase
(Kavkažani, Etiopljani, Mongoli, Ameriđani (Amerindi ili američki
starosedioci), itd.). Vidi sliku 3. Pripadnik bilo koje od ovih rasa
potencijalno bi bio međusobno-plodan sa parom suprotnog spola bilo koje
druge rase.
Nije neuobičajeno pronaći u antropološkoj
literaturi reference za do osam ljudskih geografskih rasa sa čak
dodatnim među-populacijama. Međutim, nije moja namera u ovom naučnom
radu da ulazim u diskusije različitih opcija onoga što je izraženo ovde u
tekstu ili u bilo kojim slikama, nego samo da ilustrujem navedene
taksonomske principe.
Drugi holobaramin se može sastojati od
morskih kornjača (vidi Vajz, 1992.; Robinson, 1997.). Diagram koji
pokazuje uopštene oblike živih i fosilnih kornjača može se pronaći u Luc
i Mjuzik, 1997., str. 8. Ovaj dijagram je nazvan „kladogram“ i zasnovan
je na studijama specijalista Gefnija i Mejlana, ali svi autoriteti se
ne slažu sa njihovom procenom postojećih podataka. Vidi takođe Hirajama,
1998.. Slika 4 je veoma uopšten prikaz za sve žive i izumrle morske
kornjače. U svim ovim tipovima studija pravi cilj sistematike diskontinuiteta je putem empirijskog dokaza odrediti granice zajedničkog porekla i tako ih skupiti u holobaramine.
Slika 3
Slika 4
Slika 5
Različiti pripadnici holobaramina mogu
nastati od iz-sortiranja potomstva od drugačijih gena (DNK) od
roditeljskih organizama. Ovo je uobičajeno dešavanje danas. Ili, od
vremena stvaranja mogle su biti neke nasledne modifikacije DNK
(mutacije) i ovi su prenešeni na razilažeće potomstvo. Selekcija u
prirodi je mogla uticati na potencijal za preživaljavanje raznolikog
srodstva.
Monobaramini
Drugi termin koji se koristi u baraminologiji je monobaramin (mono od grčkog jedini ili jedan). Termin monobaramin je definisan od Remajn (1993. str. 444) kao: grupa koja sadrži samo
organizme u vezi povezane zajedničkim poreklom, ali ne nužno sve od
njih. (Grupa se sastoji od jednog celog holobaramina ili njihovog dela).
Kada je holobaramin predstavljen drvetom,
jedna ili više grana tog drveta bi bila monobaramin. Na primer, među
ljudima, kavkažani bi bili monobaramin (slika 5.). Ili, za morsku
kornjaču pet trenutnih tipova koji žive u okeanu oko sveta čine
monobaramin (slika 6A od Freira, 1982.; i vidi Iverson, 1992., str. 80).
Takođe, grupa zelenih kornjača Chelonia ili grana koja sadrži ridlejsku kornjaču, Lepidochelys,
svaka bi bila monobaramin (slika 6B). Individue ili grupe mogu biti
nazvati monobaraminski ako predstavljaju delove holobaramina (slike 5,
6A, 6B i 7B).
Dodatno, sistematske studije o posebnim
monobaraminima gde je bilo divezifikacije (kao pojavljivanje
prepoznatljivijih vrsta, podvrsta, varijeteta, itd.), istraživanje na
monobaraminu ovisno o sistematskoj filozofiji istraživanja se ne bi
suštinski razlikovalo. Razlika između (1) filetskog, u smislu darvinske
makroevolucijske perpektive i (2) baraminskog (kreacionisti, „ograničene
promene“ ili mikroevolucijske, sistematika diskontinuiteta) gledišta
uglavnom bi bila da prethodni uključuje korištenje empirijskih podataka
za ekstrapoliranje nekim shvaćenim ranijim predcima. Ali, baraminolog
smatra da shvaćanje o filogeniji ne treba se proširiti izvan uverljivih
dokaza, i da se naučnici trebaju olakšati svojih osećanja obaveze
filozofski da konstruišu proširene filogenije (evoluciona drva) u
nedostatku prinudnih činjenica.
Slika 6A
Slika 6B
Apobaramini
Treći baraminski termin je apobaramin (grčki apo,
izvan od), koji „je grupa koja čini celokupnost od barem jednog
holobaramina“ (Vajz, 1999.-2000.). On može sadržavati jedan holobaramin
ili više od jednog holobaramina. „Ali mora sadržavati celokupnost od
svakog, od jednog ili više holobaramina unutar njega“. Nijedan pripadnik
organizama holobaramina unutar apobaramina ne deli poreklo sa
organizmom izvan vlastitog holobaramina (ovo je zasnovano na definiciji
holobaramina). Vidi sliku 7A.
Pridev apobaraminski se odnosi na odnos
između ili među određenih nepovezanih grupa (holobaramina). Na primer,
svi ljudi kao grupa bi bili apobaraminski, jer nijedan od pripadnika ne
deli poreklo sa bilo kojim drugim organizmima. Grupa svih ljudi i svih
kornjača takođe bi bili apobaraminski, jer nijedan čovek ili kornjača ne
dele poreklo sa bilo kojim organizmom ne-čoveka ili ne-kornjače.
Veruje se da konji (konji, magarci i
zebre) su svi povezani, jer mogu hibridizirati i stoga oni pripadaju
holobaraminu. Dodatno, postoji „pseći“ holobaramin sa monobaraminskim
granama za vukove, drugi za hijene, drugi za kojote, za šakale i više za
stotine domaćih rasa pasa. „Mačke“ čine drugi holobaramin sa
monobaraminskim grana za lava i tigra, za pume, drugi za risove, domaće
mačke, itd. (Vidi O Brajan, 1997.). Grupa svih konja (ekvidi), svih pasa
(kanidi), i svih mačaka (felidi) bi bili apobaraminski, jer nijedan
konj ili pas, ili mačka ne dele genetsku povezanost sa bilo kojim
organizmom koji nije konj, pas ili mačka.
Apobaramin kornjače može sadržavati
jedan, dva, tri ili četiri holobramina (vidi Bajz, 1992.). U ovom
trenutnom naučnom radu ja smatram da morske kornjače čine holobaramin,
stoga grupi koja čini holobaramin morske kornjače, svi ekvidi, svi
kanidi, i svi felidi bi bili apobaraminski, jer nijedan pripadnik bilo
koji od ovih četiri holobaramina ne deli genetsku povezanost sa bilo
kojim primerkom izvan njihovog dotičnog holobaramina.
Termin apobaramin je termin
koristan posebno tokom procene dva tipa organizama (parne usporedbe).
Korištenje parnih usporedbi je najuobičajenija taksonomska procedura.
Na primer, trenutni Red primata uključuje
čovekolike majmune, ljude, lemure, majmune i tarsijere. Sve rase ljudi
pripadaju jednom holobaraminu, dok šimpanze zajedno sa gorilama su
pripadnici drugog holobaramina. Tako da grupa koja sadrži ljudski
holobaramin i šimpanza-gorila holobaramin bi bila apobaraminska (vidi
sliku 7A). Dalje, skup ljudskog holobaramina sa bilo kojim ili svim drugim
primatskim holobaraminima bi bio apobaraminski. Nijedan pripadnik bilo
kojeg od ovih holobaramina bi delio bilo kakvo poreklo sa pripadnikom
bilo kojeg drugog holobaramina unutar ili čak izvan ovog apobaramina.
Vidi Robinson i Kevena, 1998. za baraminsku studiju primata.
Zaključci u vezi holobaraminskih
šimpanza-gorila odnosa (slika 7A i 7B) su zasnovani na Hartvig-Šerer,
1998.. Takođe, Kevena (1999.-2000.) me je obavestio da ponovo
istraživanje podataka od Robinsona i Kevena (1998.) podupire
holobaraminski status šimpanza i gorila. Međutim, kao što je objašnjeno
od Klajna (1999. str. 135-136) fosilni zapis daje veoma malo što je od
bilo kakve koristi u shvaćanju istorije šimpanza i gorila. Kada su
fosili uverljivo utvrđeni da su povezani sa šimpanzama i gorilama oni
trebaju biti dodani njihovom holobaraminu. Čak moguće, šimpanza-gorila
grupa treba biti podeljena u dva odvojena holobaramina. Slika 7A
ilustruje kako apobaraminska kategorija može biti korisna posebno u
slučajevima gde uključeni holobaramini sadrže primerke sa značajnom
sličnošću tokom holobaraminske granice. Tako da ljudi mogu biti
uspoređeni sa organizmima strukturno i funkcionalno najsličnijim njima,
znači šimpanzama i gorilama.
Slika 7A
Slika 7B
Polibaramini
Četvrti termin, polibaramin (poli,
od grčke reči za mnogo) je korišten za još jednu pomešanost nepovezanih
organizama. Definisan je kao grupa (dvoje ili više primeraka) koja se
sastoji delom od najmanje dva holobaramina. Može biti bilo koji od
brojnih skupova koji mogu sadržavati holobaramine, monobaramine,
apobaramine i pojedinačne primerke. (vidi sliku 8).
Kao primer, polibaramin može obuhvatati
predstavnike svih ljudskih rasa, dve vrste SAD kutijske kornjače, jednog
psa, jednog lava, jednog tigra i jednu biljku suncokreta. Ljudi čine
holobaramin. Kutijska kornjača pripada monobaraminu u kornjačinom
holobaraminu, pas različitim monobaraminima (u kanidskom holobaraminu),
lav i tigar drugom monobaraminu (u polju holobaramina) i suncokret
monobaraminu unutar biljnog holobaramina.
Pridev polibaraminski se odnosi
na odnos između ili među nekim ili svim delovima polibaramina. Na
primer, suncokret i ljudski holobaramin bi bili polibaraminski, kao što
bi bio suncokret kada se usporedi sa monobaraminom kutijske kornjače i
psom.
Ako se istraživač bavi polibaraminom
njegov taksonomski cilj trebao bi biti da razdvoji njegove delove u
druge tri kategorije što je brže moguće. Svaki od monobaramina trebao bi
biti oduzet iz polibaramina i tada pažljivo, zasebno proučavan da bi se
odredilo njegovo mesto u njihovom vlastitom holobaraminu. Pas i
suncokret svaki su deo vlastitih, različitih holobaramina i svaki od
ovih holobaramina su nepovezani bilo kojem drugom holobaraminu u ovom
polibaraminu. Takođe, ljudi trebaju biti analizovani pojedinačno da bi
se utvrdili njihovi odgovarajući odnosi unutar holobarmina.
Dalje zamisli
Da ponovimo i proširimo ovo donekle
dalje, model darvinske makroevolucije je predstavljen jednim drvetom
povezanosti, svaki oblik života je povezan sa svakim drugim oblikom
života (slika 1). U baraminskom modelu postoji šuma drveća bez povezanih
korenja (slika 2). Neki od ovih bezkorenskih drveća će imati grane
predstavljajući samo ljudsku raznolikost, drugi za kanide, drugi za
felide, itd.
Za ljude othranjene na evolucionoj dijeti
gornji meni može biti težak za progutati i svariti, jer su studenti
biologije naučeni da misle o genetskim vezama prije nego genetskim
nepovezanostima. Ali postoji manjak dokaza za povezivanje bilo koje
holobaraminske grupe za bilo koju drugu holobaraminsku grupu. Ovo je
istina ujedno za izumrle i žive tipove života.
Uobičajeno je za naučnike da koriste
drveće da opišu povezanosti, ali baraminolog Dejvid Kevena veruje da
„drveće“ moguće ne mora biti najbolji način da oslika povezanost, ali
„druge strukture kao mreže ili rešetke mogu obaviti bolji posao unutar
mnogih holobaramina. Alati prepoznavanja šablona, kao projekcijske mape
mogu moguće biti čak i bolji metodi“ (Kevena 1999.-2000.). Tako da
ostaje da vidimo upravo kako će povezanosti popularno biti predstavljene
u budućnosti.
Baraminska terminologija
Četiri termina, holobaramin, monobaramin, apobaramin i polibaramin
zvanično i javno su predstavljeni od Valter ReMina (1990. godine) na
Drugoj internacionalnoj konferenciji o kreacionizmu u Pitsburgu, PA, 30.
jula 1990. godine. Kasnije tokom nedelje iste konferencije Kurt Vajz
(1990. g.), koji je imao obilnu prepisku sa Remajn od 1983. godine,
potvrdio je Remajnovu sistematiku diskontinuiteta, venčao je sa
vlastitom mladozemljaškom kreacionističkom pozicijom i tvrdio je da je
ime ove nove sistemske procedure „baraminologija“.
Slika 8
Vajz je takođe uveo peti termin, naime arhebaramin,
koji se može shvatiti kao originalno-stvorena(e) individua(e) svakog
holobaramina. Za ljude, Adam i Eva bi činili arheobaramin. Dva druga
termina koja je Vajz uveo su neobaramin koji se odnosi na žive oblike života i paleobaramin
za starije organizme. Arheobaramini su najteoretičniji (Vajz, 1990.);
dok holo-, mono-, apo-, paleo- i neobaramini se mogu odrediti na osnovi
posmatrane informacije.
Baraminologija na delu
Važno je naglasiti da striktno empirijska
komponenta baraminologije je sistematika diskontinuiteta koja može biti
korištena sama za sebe bez ikakvog pozivanja ka religijskoj
litetaraturi. Zapravo, većina sekcija u predstavljenom sadašnjem naučnom
radu uključujući slike su zapravo zasnovani na nepovezanostima
promatranim u prirodi. Remajn (2000. g.) je pokazao da je sistematika
diskontinuiteta namerno dizajnirana da bude neutralna, naučni metod za
proučavanje nekih od naučnih šablona. Mi ne započinjemo
pretpostavljanjem nepovezanosti, nego mi sledimo podatke da identifikujemo
nepovezanost, gde god bi mogla biti. Ovaj sistematski metod je
empirijski, naučni poduhvat-pokretan podacima, a ne teoretskim
pretpostavkama.
U stvarnom procesu kretanju prema cilju
prepoznavanja holobaramina, taksonom treba idetifikovati apobaramine i
podeliti ih. Oduzimajući kriterij treba biti korišten u deljenju
apobaramina u izdvojene holobaramine. Tada, sa ciljem karakterisanja
holobaramina, taksonom se usredsređuje na monobaramine i dodatni
kriterij je korišten da izgradi ove monobaramine.
Analogija za objašnjenje ovog procesa je predložena:
To je kao da je bio ogroman pad snega
koji je prekrio drveće do vrha, i mi kopamo dole u sneg da
identifikujemo povezanosti, grane, ogranke i deblo. Da li je jedno
drvo ispod? Ili je to voćnjak odvojenih, različitih drveća? Dok podaci
polako dolaze u vidokrug mi ćemo imati argumente šta je povezano s čim,
ili da li postoji nepovezanost na danom mestu. Neki istraživači će
pogrešno identifkovati različito granje kao da je povezano, dok su ovi
poslednji nepovezani i tako dalje. Ali ovo se otkriva dok kopamo. Mi ne
„sečemo i ne obrezujemo“ podatke. Radije, mi ostavljamo podatke upravo
gde su. Mi samo sečemo i obrezujemo naše percepcije, osobito naše
privremeno pogrešne percepcije podataka (Remajn, 2000.).
Drugim rečima, naučnici ponavljaju opitne
taksonomije povećavanjem ili smanjivanjem veličine grana da dostignu na
najbolju približnost realnosti. Ova sistematska procedura je vođena
promatranim činjenicama prije nego nekom pretpostavljenom konstrukcijom.
Cilj baraminologije je prepoznati
holobaramine, ali baraminolozi ne prepoznaju holobaramine kao apsolutno
različite od bilo apobaramina ili monobaramina. Apobaramini sadrže jedan
ili više holobaramina. Tako da ako je apobaramin razdeljen i samo jedan
holobaramin je ostao da li je taj holobaramin i dalje apobaramin?
Baraminolog kaže da jeste.
Takođe, ako postoji deo (grana)
holobaramina ona je nazvana monobaramin. Monobaramin će rasti u veličinu
i kompleksnost kako su više primeraka grana dodani. Kada je taksonom
dodao sve grane koje mogu biti nađene među trenutno živim ili izumrlim
organizmima, takonom može presuditi da je drvo kompletno prema svim
trenutno postojećim i primenjivim informacijama. Ovo znači da „drvo“,
koje može imati jednu ili više grana, ima sve oblike života za koji se
veruje da deli genetsku povezanost, što znači da su oni povezani
poreklom. Tako da ova grupa sada po definiciji će biti holobaramin. Da
li je i dalje monobaramin? Baraminolog kaže da! Onda kako može grupa
biti monobaramin ili apobaramin i biti holobaramin u isto vreme?
Slika 9
Remajn (1993., str. 447) i Vajz
(1999.-2000.) objašnjavaju da mi možemo misliti u terminima postavljene
teorije. Uzmite u obzir veliki krug, A (apobaramin). Unutrašnji deo toga
je manji krug B (holobaramin), i ovo uključuje još manju unutrašnju
regiju C (monobaramin). Vidi sliku 9. Ujedno apobramin (A) i monobaramin
(C) su promenjeni u smeru srednjeg kruga, holobaramina (B).
Smernice
U postizanju cilja odvajanja delova
polibaramina, deljenja apobaramina, izgradnje monobaramina i
prepoznavanju holobaramina, taksonom treba smernice za odlučivanje šta
pripada osobitoj monobaraminskoj grani. Ovi standardi će varirati ovisno
o grupama koje se razmatraju, ali opšte smernice koje su upotrebljavaju
uključuju:
- Tvrdnje Svetog Pisma (korištene u baraminologiji ali ne u sistematici diskontinuiteta). Ono ima prioritet nad svim drugim razmatranjima. Na primer, ljudi su odvojeni holobaramini, jer su odvojeno stvoreni (Postanje 1 i 2). Međutim, čak i kako je objašnjeno od Vajza u njegovoj od 1990. godine govornoj prezentaciji, ne postoje veoma bitne taksonomske informacije u Bibliji. Takođe, Remajnova sistematika diskontinuiteta, jer je neutralan naučni poduhvat, ne uključuje Bibliju kao izvor taksonomske informacije.
- Hibridizacija. Istorijski, Marš i drugi su postavili ovaj kriterij sekundaran jedino Bibliji, jer ako plodan potomak može biti dobijen od ukrštanja između dva različita oblika, ovo bi definisalo njihov monobaraminski status. Međutim, mi shvaćamo danas da manjak znane hibridizacije između pripadnika različitih populacija organizama ne nužno samo po sebi znači da nisu u rodu. Hibridizacijski kriterij verovatno će zadržati validnost, ali je ponovo razmatran u svetlu moderne genetike.
- Ontogenija, naime razvoj individue od embrija do odrasle jedinke. Hartvig-Šerer (1998.) sugerira da komparativna ontogenija sledi hibridizaciju u važnosti kao kriterij za pripadnost osobitom tipu.
- Loza. Da li postoji dokaz jasno očite loze između i među bilo, ili oboje fosilnih i živih oblika.
- Struktura (morfologija) i fiziologija (funkcija). Strukture mogu biti makroskopske (veliki entiteti kao telesni organi), mikroskopske (mali, i promatrani povećavanjem), i molekularne (hemijski) konfiguracije.
- Fosili u slojevima stene. Ove studije mogu uključiti lokacije fosilnih oblika u slojevima stene, i mogu povlačiti sa sobom razmatranje sedimenata Potopa.
- Ekologija. Važno je shvatiti utočište organizma, to jest regiju gde živi i kakvu ima interakciju sa okolinom uključujući druge žive stvari.
Da bi utvrdili baraminska rastojanja među
tipovima organizama važno je da iskoristimo najvažnije podatke. Na
primer, molekularne studije sa mitohondrijskim DNK i RNK su bile korisne
sa nekim kornjačama, ali autor je doveo u pitanje baraminsku korisnost
ekoloških kriterija (Robinson, 1997.). U baraminskom istraživanju ljudi
sa ne-ljudskim primatima, morfološka (oblici) obeležja kao zubi i kosti
kao i ekološke osobine uključujući hranjenje i postojbine su bile puno
vrednije od hromozomskih ili molekularnih (hemoglobin i RNK) informacija
(Robinson i Kevena, 1998a.). Takođe, vidi Garsia-Pozuelo-Ramos, 1997.;
1998.; 1999.. Međutim, baraminsko istraživanje na širem spektru felida
je otkrilo da ekološki podatci su najmanje verodostojni, i hromozomskii
podaci male pouzdanosti. Morfološke i molekularne (protein i RNK)
informacije su bile najvažnije (Robinson i Kevena, 1998b.). Za daljnje
studije Cavan (1999.-2000.) je nedavno naglasio da:
Osobito, proteini i njihov DNK šablon,
koji su delovi fundamentalnih ćelijskih procesa, imaju široku
primenjivost u baraminološkom istraživanju. Na primer, proteini koji su
udruženi sa celularnom respiracijom kao citohrom C i citohrom B su
odlični kandidati. Vidi na primer, citohrom C tablicu u Dentonu (1986.).
Statistika koja uključuje kompjuterske
proračune su bitni u daljnjim istraživanjima. Treba biti očekivano da
kada je barminologija široko prihvaćena nauka taksonomije će biti
revolucionizirana. Kada sistematici rade sa „šumom“ drveća prije nego
jednim velikim evolucionim drvetom moguće je da kategorije kao
kraljevstva, filuma, podela, klasa i čak redovi će biti manje korisni u
klasifikaciji. Međutim, među živim stvarima, grupe obeležja unutar
drugih grupa ili karakteristike mogu biti promatrane. Ovi nazvani
„ugnežđeni šabloni“ (vidi Remajn, 1993.; Vajz 1998.) mogu se proširiti
iznad baraminskih kategorija, tako da fenetske i kladističke metode mogu
nastaviti da budu korisni uz sistematiku diskontinuiteta.
Za one koji su ogrezli u Linejsku
taksonomiju i evoluciono razmišljanje, sistematika diskontinuiteta može
izgledati kao besmislen predlog. Međutim, kao što je poznato smeona šema
ne treba biti shvaćena kao odstupanje od realnosti. Zanimljivo, prvog
avgusta tokom 1999. godine na Internacionalnom kongresu botanike u Sent
Luisu preplavljena gomila je čula prezentaciju koja promoviše tzv.
FiloKod, sistematsku šemu koja će dovesti do ukidanja kraljevstava,
filuma, klasa, redova itd. (Milius, 1999.). Takođe vidi naučne radove od
de Kviroza (1992.; 1997a.; 1997b.). Predložena nekomplicirana
sistematska procedura se usredsređuje na kladuse, svaki kladus se
sastoji od jedinstvene vrste i potomaka od te vrste. Drugim rečima,
kladus će biti holofilska (genetski-ujedinjena) grupa.
Međutim, oni koji koriste šemu kao ovu
uopšteno misle o kladusu unutar kladusa unutar većih kladusa na
makroevolucionoj skali, dok baraminologija je više mikroevoluciona (male
promene) koja je manje spekulativna. Za drugu taksonomsku literaturu
koja daje potporu tipologiji vidi Šerer; 1993..
Koliko će mnogo holobaramina biti –
3.000, 5.000, 10.000, 15.000, ili više? U ovo vreme najbolji ogledni
odgovor je „verovatno nekoliko hiljada“.
Aktivni baraminolozi
Dodatno gore spomenutih osam ljudi, bili
su tu takođe Pit Viliams, Džeri Kreps i Džef Tompkins koji su postali
umešani u različitom stepenu. Oktobra 1998. godine Džoi Daso i ja smo
prihvatili pozive da se pridružimo grupi, tako povećavajući broj članova
na 13.
Tri dana konferencije sa povoljnim
naslovom „Baraminologija 99: Stvaranje biologije za 21. vek“ je
organizovano i predstavljeno od BSG u saradnji sa Departmenima biologije
i hemije na Liberti univerzitetu u Linčburgu, VA, 5.-7. avgusta 1999.
godine. Bilo je 24 pozvanih učesnika.
Formalna prezentacija je napravljena od
Tod S. Vuda, D. Ešli Robinsona, Kurt P. Vajza, Pit Dž. Viliamsa, i Pol
Nelsona. Teme koje su se odnosile na baraminologiju su uključivale
kreacionističku biologiju, filogenesku inferenciju, biblijske studije,
teoriju dizajna, hibridizacijski kriterij, evolucione mreže i
ne-sistematska polja. Zadnje posle-podne je posvećeno radionici o
baraminologiji porodice koja se sastoji od kamila. Ove prezentacije i
diskusije o kamilama su uključivale uvod, biblijske i lingvističke
studije, fosilni zapis, hibridizaciju, molekularne studije, filogenske
distorcije i zaključke. Zadnje veče konferencije je uključivalo umerenu
diskusiju o „Budućnosti BSG i baraminologije.“
Razmatranje je obavljeno o uspostavljanju
novog duštva i novog časopisa. Učesnici su volontirali o odgovornosti
saopštavanja na drugoj konferenciji na zapadnoj obali na početku 2001.
godine.
Bez obzira na neke razlike u mišljenima,
moj zaključak je bio da je grupa kao celina bila veoma uzbuđena
napretkom koji je postignut do sada i perspektivama budućnosti
baraminologije. Neki od naučnika su preferirali da veruju u staru zemlju
(milijarde godina), ali vođstvo i većina prisutnih su bili ujedinjeni u
održavanju perspektive mlade zemlje (hiljade godine starosti). Uvod u
braminologiju i izveštaj sa konferencijie od 1999. godine su napisani od
prisutne, botaniste Margaret Helder, 1999. godine.
Zaključni komentari
Baraminologija može biti smatrana kao
tipološki pristup klasifikaciji oblika života, živih i fosilizovanih. U
prošlim vekovima naučnici su obično tipološki više teoretisali nego što
čine sada. Međutim, zbog mnogih teškoća (na primer, konvergencije i
inverzije) koji je mučio makroevolucione mislioce, postoji rastuća
prijemljivost za tipologiju.
Baraminolozi veruju da su oni u prvom
redu modernog progresivnog mišljenja. Oni zainteresovani za učenje više o
deset godina starom polju baraminologije trebalo bi da razmotre
reference.
Zasluge
Dejvid Kevena, Kurt Vajz i Valter Remajn
su pročitali ranije skice ovog rukopisa i doprineli sa puno vrednih
informacija. Druga pomoć je došla od Džon V. Kuoza, Nikl Dorana, Skot A.
Mateja, Tod Vuda i D. Ešli Robinsona. Pomoć u mehaničkim detaljima je
obavljen od Lejn P. Lestera, CRSQ Managing editor. Naučni rad je napisan kao reakcija na strpljivo poticanje od CRSQ editora, Emet L. Viliamsa.
Reference
CRSQ: Creation Research Society Quarterly
Bartz, Paul A. 1991. A refinement of biosystematics which reflects baraminic variation. CRSQ 28(1):18–20.
Cavanaugh, David P. 1999–2000. Personal communications.
Cuozzo, Jack. 1998. Buried alive: the startling truth about Neanderthal Man. Master Books, PO Box 727, Green Forest, AR 72638.
. 2000. Personal communications.
Denton, Michael. 1986. Evolution: a theory in crisis. Woodbine House, Rockville, MD.
de Queiroz, Kevin. 1992. Phylogenetic taxonomy. Annual Review of Ecology and Systematics 23:449–480.
. 1997a. Misunderstandings about the phylogenetic approach to biological nomenclature: a reply to Liden and Oxelman. Zoologica Scripta 26(1):67–70.
. 1997b. The Linnaean hierarchy and the evolutionization of taxonomy, with emphasis on the problem of nomenclature. Aliso (The Rancho Santa Ana Botanic Garden, Claremont, CA 91711–3157) 15(2):125–144.
Frair, Wayne. 1982. Serum electrophoresis and sea turtle classification. Comparative Biochemistry and Physiology 72B:1–4.
. 1991. Original kinds and turtle phylogeny. CRSQ 28(1):21– 24.
. 1999. Creationist classification—an update. Creation Matters 4(1):1,5.
Garcia-Pozuelo-Ramos, Celedonio. 1997. Taxonomy of primates. CRSQ 34(2):93–102.
. 1998. Dental variability in the domestic dog (Canis familiaris): implications for the variability of primates. CRSQ 35(2):66–75.
. 1999. Craniodental variability in the domestic dog (Canis familiaris) and its implications for the variability in primates. CRSQ 36(3):116–123.
Hartwig-Scherer, Sigrid. 1998. Apes or
ancestors?: interpretations of the hominid fossil record within
evolutionary and basic type biology. In William A. Dembski, editor. Mere creation: science, faith and intelligent design, pp. 212–235. Intervarsity Press, Downers Grove, IL.
Helder, Margaret. 1999. Introducing baramins. Creation Dialogue (5328 Calgary Trail South, Suite 1136, Edmonton, Alberta T6H 4J8) 26(3):6–7.
Hirayama, Ren. 1998. Oldest known sea turtle. Nature 392:705–708.
Iverson, John B. 1992. A revised checklist with distribution maps of the turtles of the world. Privately printed, Richmond, IN.
Klein, Richard G. 1999. The human career : human biological and cultural origins. The University of Chicago Press, Chicago, IL.
Lutz, Peter L., and John A. Musick. 1997. The biology of sea turtles. CRC Press, New York.
Marsh, Frank L. 1941. Fundamental biology. Published by the author, Lincoln, NE.
. 1969. The form and structure of living things. CRSQ 6(1):13–25.
Milius, Susan. 1999. Should we junk Linnaeus? Science News 156(17):268–270.
O’Brien, Stephen J. 1997 (June). The human-cat connection. National Geographic 191(6):77–85.
Remajn, Walter J. 1990. Discontinuity
systematics: a new methodology of biosystematics relevant to the
creation model. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 207–216. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
. 1993. The biotic message: evolution versus message theory. St. Paul Science, PO Box 28006, Saint Paul, MN 55128.
. 2000. Personal communications.
Robinson, D. Ashley. 1997. A mitochondrial DNA analysis of the testudine apobaramin. CRSQ 33(4):262–272.
Robinson, D. Ashley, and David P.
Cavanaugh. 1998a. A quantitative approach to baraminology with examples
from the catarrhine primates. CRSQ 34(4):196–208.
. 1998b. Evidence for a holobaraminic origin of the cats. CRSQ 35(1):2–14.
Scherer, Siegfried. 1993. Typen des Lebens. Pascal Verlag, Berlin.
Williams, Pete J. 1997. What does min mean? Creation Ex Nihilo Technical Journal 11(3):344–352.
Wise, Kurt P. 1990. Baraminology: a young-earth creation biosystematic method. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345–360. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
. 1992. Practical baraminology. Creation Ex Nihilo Technical Journal 6(2):122–137.
. 1998. Is life singularly nested or not? In R. E. Walsh, editor, Proceedings of the Fourth International Conference on Creationism. Technical Symposium Sessions, pp. 619–631. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
. 1999–2000. Lične komunikacije.
Pretplati se na:
Postovi (Atom)